

バイオサイトジェンのゲノム編集プラットフォームは2009年の会社設立とともに設置されました。これは主として動物、細胞モデルの研究開発を目的としています。ゲノム編集モデルは基礎科学研究において大切な役割を果たすだけでなく、抗体医薬品開発過程における多くの重要な問題を解決するものとなります。完全ヒト抗体産生マウスRenMabを使用することで、完全ヒト配列に基づきマウスの免疫システムがスクリーニングした抗体分子を直接取得することができます。重鎖抗体産生マウスRenNanoは、完全ヒト配列のナノ抗体の開発に用いることができます。
スクリーニングした抗体分子がヒトの創薬ターゲットだけを識別し、マウスの同種組織創薬ターゲットを識別しない場合、創薬標的ヒト化マウス・細胞モデルは、医薬品の安全性評価、薬効評価、メカニズム研究などの問題を解決することになります。上記の応用方法に加え、ゲノム編集モデルはさらに広い分野で応用が可能です。例えば、一部の遺伝子修飾による抗原の免疫反応の向上、もしくは遺伝子の変異誘導モデルによる疾患モデルの作製などに活用できます。
多年にわたる技術開発の結果、3つの主要なゲノム編集技術体系が構築されることになりました。(1)マウスの胚性幹細胞に基づいたESC/HR技術。私たちが確立したマウス胚性幹細胞株は約70世代にわたり分化全能性を保持しているため、同一染色体で何回もの遺伝子改造を行うことができます。(2)EGE技術。CRISPR/Cas9が媒介する遺伝子ノックイン効率を約20倍向上させることができます。(3)SUPCE技術。DNAセグメントの長さによる制限を受けず、最高3回のマウス胚性幹細胞操作を通し、一回で超大型遺伝子セグメントをヒト化することができます。この3種類の技術体系のおかげで、私たちはゲノム編集技術に存在する問題を解決し、高効率で大規模の多重改造や超大型遺伝子のヒト化などを実現することができます。セグメントの長さや部位にかかわりなく、正確なゲノム編集が行えるようになったのです。
ESC/HR
ESC/HR (ESC, Embryonic Stem Cell; HR, Homologous Recombination) とは、ドナーDNAとゲノム間の相同組換えを利用してマウスの胚性幹細胞のゲノム編集を行う技術です。遺伝子改造の陽性クローンをマウスの胞胚に注入し、それを育種することで最終的に成体マウスを得ます。バイオサイトジェンが構築したC57BL/6ベースの幹細胞株と培養体系は、約100世代にわたり分化全能性が保証されており、適切な成体マウスが提供されます。

EGE
EGE (Extreme Genome Editing system)はCRISPR/Cas9をベースにした開発と改善を経て、遺伝子ノックインの効率を約20倍にしたゲノム編集技術です。CRISPR/Cas9は相同組換えの発生確率を大幅に引き上げ、ゲノム編集を簡単にしましたが、大規模のモデル作製においては効率が低く、コストが高いという問題が存在していました。EGEシステムはゲノム編集をさらに高速化し、迅速かつ正確にどの部位のゲノムのDNA配列でも編集できるようにしました。ゲノム編集の効率が高まると様々なメリットがあります。まず、モデル生物の作製の成功率が上がり、コスト節約になります。また時間が短縮されるため、工業化や大規模なビジネス応用が可能になります。さらに、遺伝子改造の難度はそれぞれ異なりますが、一部の難度が非常に高い部位のゲノム改造が容易になりました。このようにEGE技術により効率向上は不可能だった事柄も可能に変えたのです。
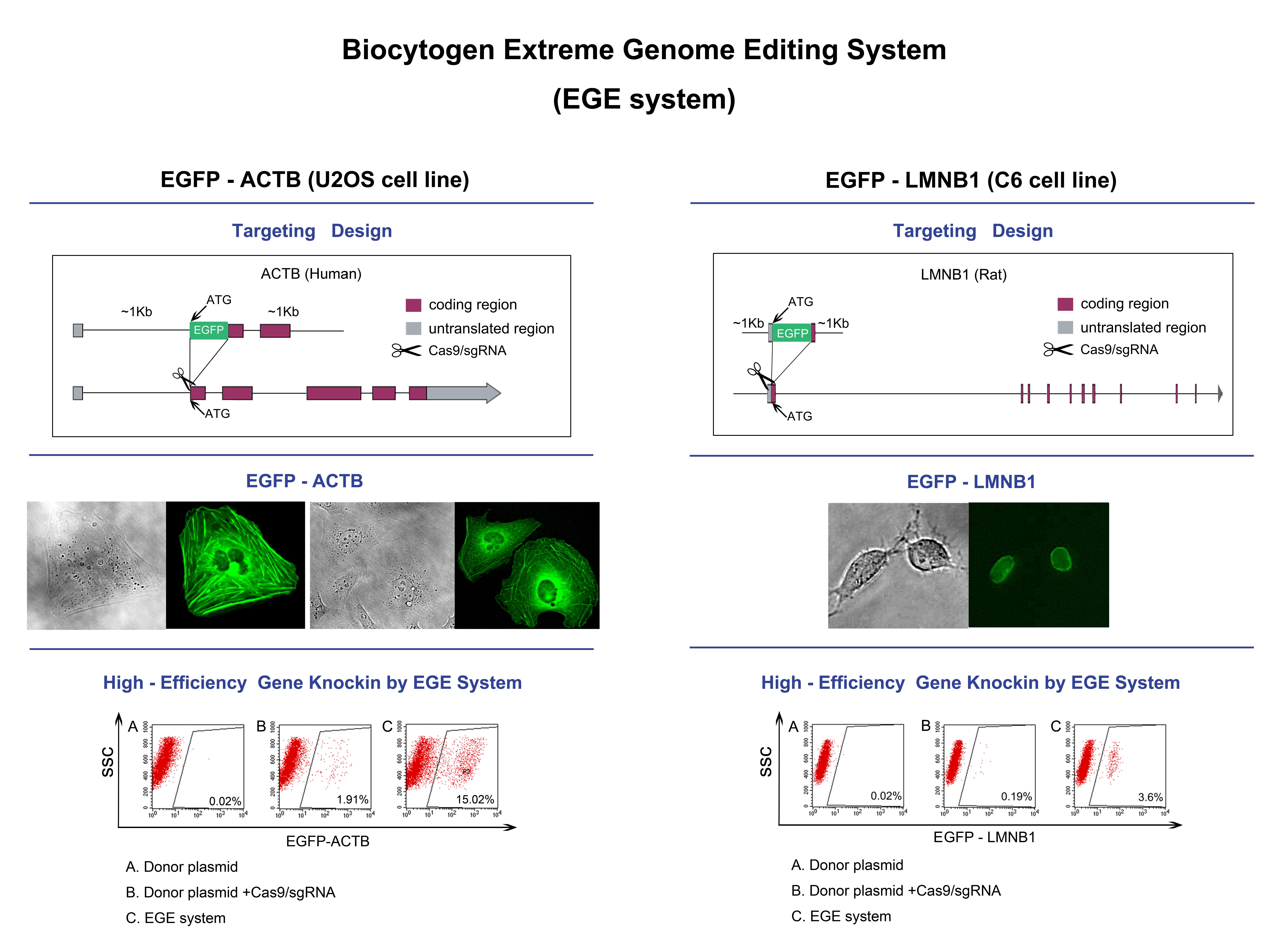
図1. EGEシステムはCRISPR/Cas9ベースの遺伝子ノックインの効率を大幅に向上させた
最初のモデル(図左)は、ヒト骨肉腫細胞を使用し、設計案をヒト細胞フレームワークタンパク質β-Actin遺伝子ACTBの開始コドンATGとした後、緑色蛍光タンパク質EGFPをノックインすると、ノックインが成功した細胞はEGFP融合β-Actinタンパク質を発現した。U2OSトランスフェクションではCas9ヌクレアーゼと標的ACTBのsgRNAプラスミドを発現し、両側には約1kbの長さのホモロジーアームのEGFPドナープラスミドが存在した。トランスフェクション後フローサイトメトリーで分析すると、基本的なCas9/sgRNA媒介のノックイン効率は1.91%だが、EGEシステムではノックイン効率を15.02%に向上した。2番目のモデル(図右)では、ラットのグリオーマC6細胞と既知コード核膜タンパク質のラットのLmnb1遺伝子をターゲットとした。このモデルではEGEシステムがノックイン効率を0.19%から3.6%にした。
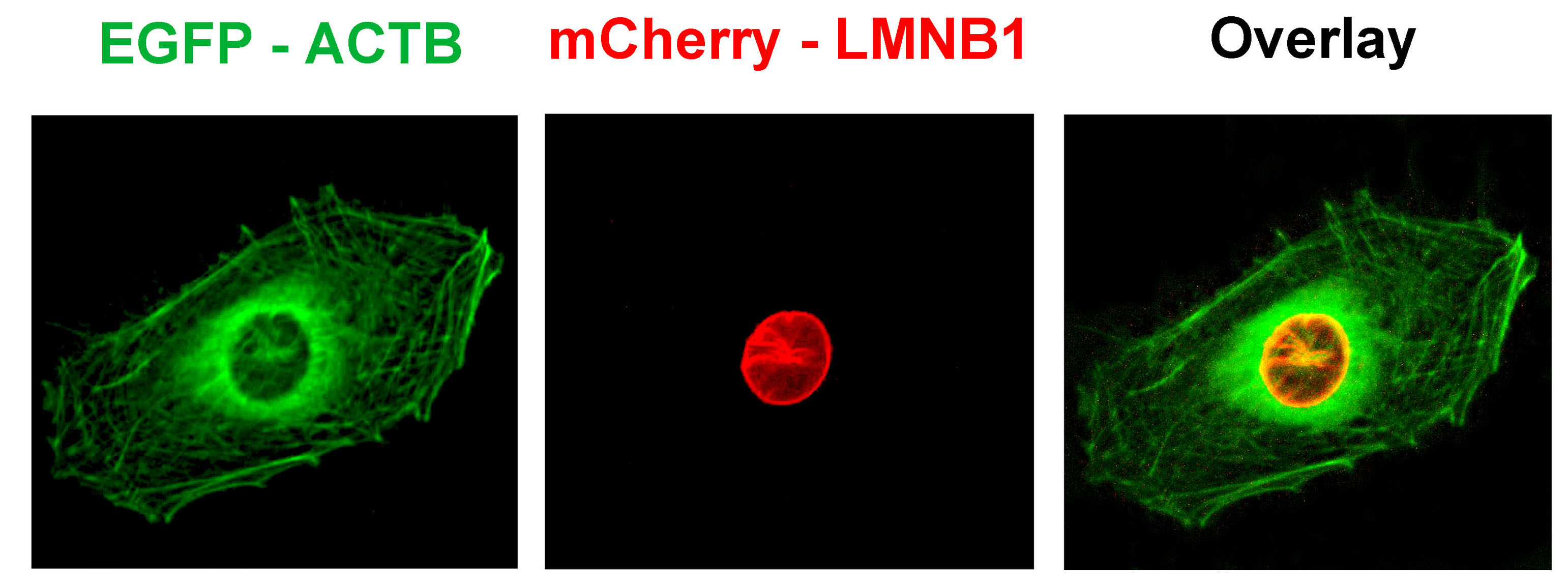
-
図2. EGE技術におり細胞中に同時に2つの遺伝子をマーキングできるようになった
U2OS細胞でACTBおよびLMNB1遺伝子をターゲットとするCas9/sgRNAプラスミド、ACTBをマーキングする緑色蛍光タンパク質GFPドナープラスミド、LMNB1遺伝子をマーキングする赤色蛍光タンパク質mCherryドナープラスミドのトランスフェクションを行った。トランスフェクション後、蛍光顕微鏡で観察したところ、赤色の核膜タンパク質と緑色の細胞フレームワークタンパク質が観察できた。
SUPCE
SUPCE (Size-Unlimited and Precise Chromosome Engineering system) はDNAセグメントの長さの制限を受けないゲノム編集技術です。これまではプラスミドまたは細菌人工染色体(BAC, Bacterial Artificial Chromosome) を使用し大型セグメントの変換のためにはマウスの胚性幹細胞を何回も改造する必要がありました。ドナー容量の制限があるため、改造の回数と変換するセグメントの長さは比例していました。マウスのES細胞はin vitroの長期操作ではその全能性を失いやすく、成体マウスが得られないこともしばしば生じました。SUPCEはこの問題を根本的に解決しました。
バイオサイトジェンはSUPCE技術を使用して完全ヒト抗体産生マウスRenMab (図1)の開発に成功しました。図2のFISH測定結果は、ヒト重鎖、κ軽鎖配列を、マウスのゲノムの対応位置へ挿入し変換に成功したことを検証するものです。SUPCE技術は、抗体遺伝子だけでなく、他の超大型遺伝子クラスターのヒト化に応用できます。例:T細胞受容体 (TCR, T-Cell Receptor)、主要組織適合性複合体 (MHC, Major Histocompatibility Complex)、NK細胞遺伝子断片のクラスター(NKC, NK Cluster) など。
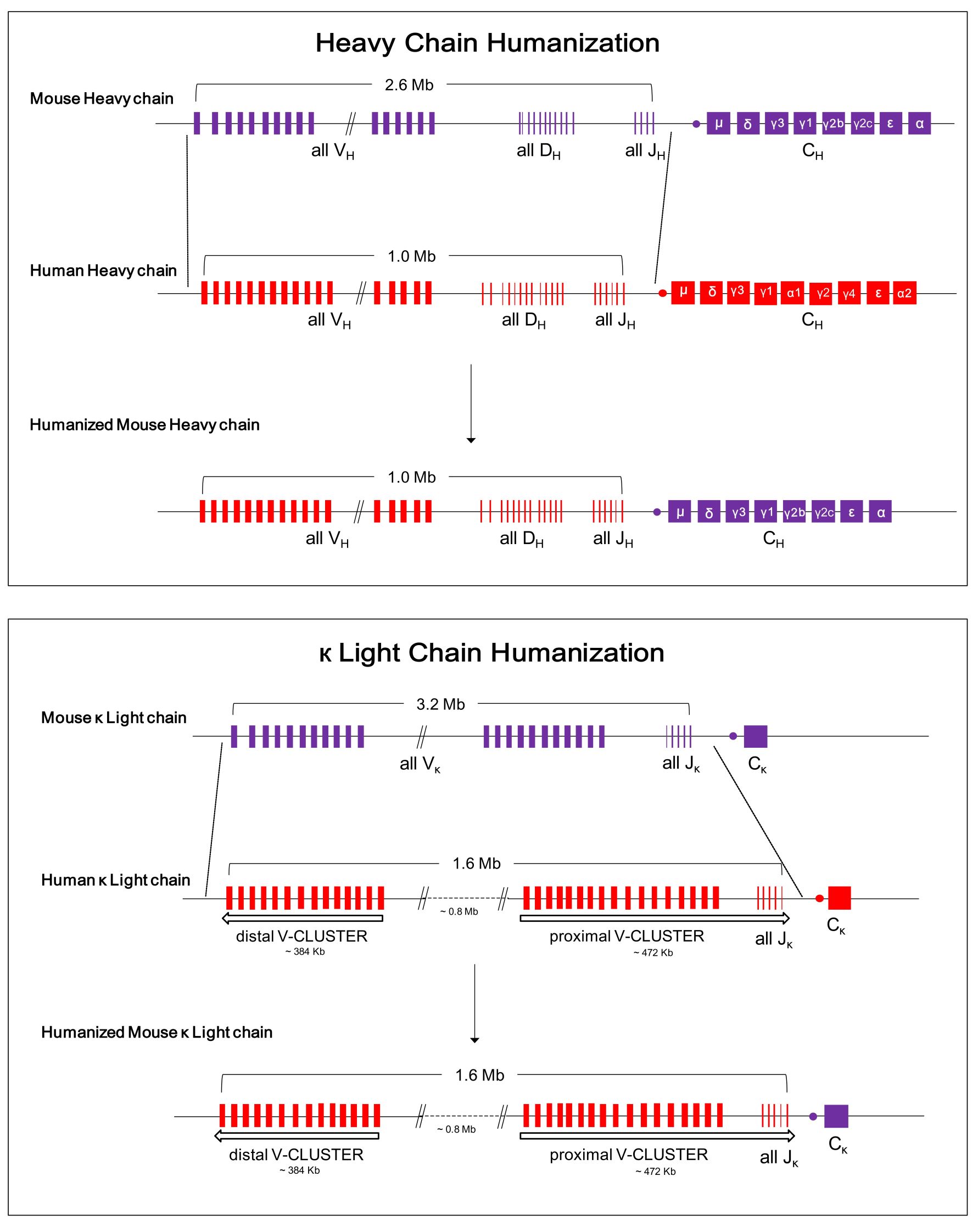
-
図1. RenMabマウスの抗体遺伝子のヒト化設計図
-
-
RenMabマウスを使用し、1回で2.6 Mbのマウス重鎖可変領域を1.0 Mbのヒト重鎖可変領域の配列に変換し、3.2 Mbのマウスκ軽鎖可変領域を1.6 Mbのヒト軽鎖可変領域の配列に変換した。
-
-
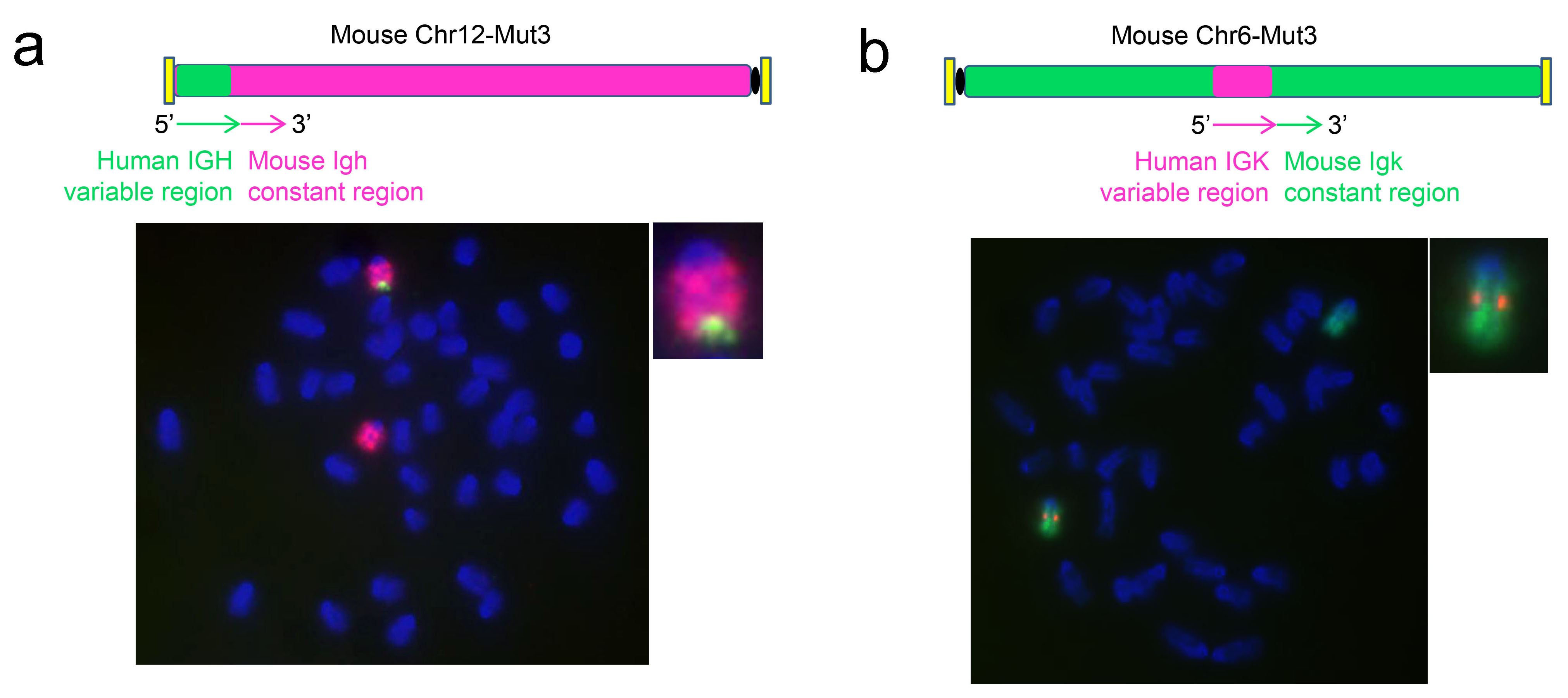
図2. 抗体遺伝子可変領域ヒト化変換のFISH測定結果
SUPCE技術を利用し、マウス胚性幹細胞のマウス重鎖およびκ軽鎖の可変領域をin situで完全なヒト由来配列に変換した。a. ヒト重鎖可変領域配列プローブ(緑色)とマウスIghがある#12号染色体全染色プローブ(赤色)で、重鎖ヒト化細胞クローンを測定。b. ヒトκ軽鎖可変領域配列プローブ(赤色)とマウスIgkがある#6号染色体全染色プローブ(緑色)でκ軽鎖ヒト化細胞クローンを測定。
自社開発によるゲノム編集技術に加え、自動化されたハイスループット技術プロセスも高速で安定した大規模モデルの作製を保証するものとなります。プラットフォームでは複数の96経路核酸抽出装置を用いてマウス・細胞ゲノムDNAとプラスミドDNAの抽出を行っています。また自動分注システムは遺伝子測定の全自動化を実現しました。ゲノム編集プラットフォームはこうして毎年1500以上のモデル開発を行っています。







